|
A. Introduction Haemopoiesis is regulated by a variety of factors determining proliferation, differentiation, amplification and maturation of stem cells and their committed progeny, the myeloid and lymphoid precursor cells. Investigation of putative regulatory molecules has been facilitated by the development of clonogenic in vitro systems whereby restricted progenitor cells are induced to undergo clonal expansion and maturation in soft gel media, resulting in discrete colonies of mature cells. Using these systems, most committed precursor cells can be grown in culture (Reviewed in Metcalf 1977). Furthermore, haemopoietic stem cells can now be maintained in vitro for many months, continuously generating myeloid and lymphoid restricted cells (Dexter et al. 1977 a; Dexter et al. 1978). In all aspects so far studied the haemopoietic cells produced in such longterm culture are apparently normal and possess characteristics in common with their counterparts present in freshly isolated bone marrow (Dexter et al. to be published a). Using these cultures we have heen investigating the regulation of stem cell proliferation and differentiation and the effects of a variety of RNA C-type leukaemia viruses on in vitro haemopoiesis (Dexter et al. 1977b; Teich and Dexter 1979; Teich et al. 1979; Dexter and Teich 1979; Testa et al.1980).
Several cell lines have been isolated from long-term cultures
infected with Friend leukaemia virus (FLY) : 1. In one experiment,
cells isolated from long-term cultures 14 days after FY infection
showed GM-CSF independent colony growth in soft agar. When individual
colonies were isolated, it was found that the cells grew antonomously
in suspension and that injection of the cells in vivo produced a
rapidly progressing myelomonocytic leukaemia (Testa et al. 1980).
This cell line, designated 427E, was aneuploid with a mean of 78
chromosomes and a constitutive producer of GM-CSF. When plated in
soft agar, colony formation occurred in the absence of added stimulatory
molecules. However, if the cells were plated in the presence of
excess exogenous GM-CSF, it was found that although the initial
colony forming efficiency was not altered, the self-renewal ability
of the colony forming cells (measured by re-plating ability) was
dramatically reduced. These leukaemia cells, therefore, show at
least some biological response when cultured in the presence of
excess GM-CSF -a proposed regulator of granulopiesis. Similar effects
of GM-CSF have been observed in other myelomonocytic leukaemia cell
lines (Metcalf et al. 1969; Ichikawa 1969; Fibach et al. 1972).
Infection of marrow culture with FBJ osteosascoma virus has similarly
led to the rapid emergence of a malignant myelomonocytic clone of
cells ( 426-C) with characteristics similar to those described above.
2. Long-term cultures treated with Abelson leukaemia virus readily
undergo malignant transformation to produce poorly differentiated
B-cell leukaemia cell lines (Teich et al. 1979; Teich and Dexter
1978). The cells grow autonomously in suspension and in soft agar
and are inducible for intracytoplasmic IgM production by various
reagents. 3. The infection of marrow cultures with FLY can also
lead to the production of apparently normal, non-leukaemic cell
lines which possess characteristics of either stem cells or committed
granulocyte progenitor cells (Dexter et al. 1979; Dexter and Teich
1979; Dexter et al. 197 8Ob ) .These celllines are characteristically
isolated only from long-term cultures which have been maintained
for several months. 416B cells were isolated from a culture more
than 5 months after infection with FLY and were established as a
continuous cell line, growing in suspension independently of added
stimulatory molecules. Upon isolation the cells were initially bipotential
(Dexter et al. 1979) when injected into irradiated mice and formed
spleen colonies containing granulocytes and megakayocytes, although
they grew in suspension (in vitro) as an undifferentiated cell population.
The cells had a normal diploid karyotype and were non-leukaemic.
Colony formation in soft agar was only seen in the presence of exogenous
GM -CSF (Dexter et al. 1978a). Eventually, karyotype instability
was seen, the cells became restricted to erythroid development when
injected in vivo and colony formation in soft agar occurred in the
absence of added GM-CSF. However, the cells were still non-leukaemic.
Another cell line, 458C, was isolated more than 5 months after FLY
infection (Dexter et al. 198Ob ), also grew autonomously in suspension
culture, maintained a diploid karyotype and was non-leukaemic. Colony
formation in soft agar initially occurred only in the presence of
added GM-CSF, and the colonies produced consisted of neutrophil
granulocytes. Presently, this cell line also has aquired the ability
to undergo clonal expansion in soft agar in the absence of GM-CSF,
and a karyotype investigation is in progress. 4. It has recently
been reported by Greenberger et al. (1979) that infection of susceptible
long-term bone marrow cultures with FLY is followed consistently
by the generation of promyelocytic leukaemia cell lines. For their
continued growth such cells must be sub-cultured in medium conditioned
by the growth of WEHI-3CM. Since one component ofWEHI3CM is GM-CSF,
it was assumed that this moiety acted as the growth promoter for
the proliferation of these cells (Greenberger et al. 1979). According
to this report, WEHI-3CM dependent cell lines could not be obtained
from control (non-infected) cultures. However in recent work we
have shown that cells from uninfected long-term marrow cultures
will consistently generate cell lines in the presence of either
WEHI-3CM or pokeweed-mitogen spleen cell conditioned medium (Dexter
et al. 198Oc). Such cell lines are non-leukaemic, maintain a diploid
karyotype and from colonies containing. granulocytes when plated
in soft agar. Growth in suspension culture or in soft agar is absolutely
dependent upon the continued presence of WEHI-3CM or SCM. Other
GM-CSF containing conditioned media or highly purified GM-CSF preparation
did not support the growth of these cells (Dexter et al. 198Oc).
This suggests that GM-CSF is not the regulatory molecule involved
in the maintenance of proliferation. We have further suggested that
such cells represent a population of committed granulocyte progenitor
cells which are capable of extensive self renewal and which are
responding to a hitherto unrecognised regulator. Thus far cell lines
have been produced from marrow cultures of strain DBA/2, C57BL/6,
BDF1 and Swiss mice. No evidence of viral replication can be found
in these cell lines, which are designated FactorDependent Continuous
cell lines, Paterson Labs (FDC-P). A summary of these cell lines
is given in Table 1. FDC-P lines produced from untreated cultures
demonstrate factor dependency for growth in suspension and in soft
agar . Initial isolates of 416B and 458C showed independent growth
in suspension but dependent growth in soft agar. These cell lines
are characterised by being diploid and non-leukaemic and apparently
undergo normal differentiation. Karyotype changes occurring in 416B
are associated with a restriction in development potential and acquisition
of factor-independent growth in soft agar . Other cell lines produced
from virus infected long-term cultures show a restricted developmental
potential, are aneuploid, leukaemic and are independent of the addition
of exogenous factors for growth in suspension or in soft agar. The
MI cells represent a line derived from cultures treated with the
carcinogen methylnitrosourea (MNU). These cells are also factor
independent, aneuploid and leukaemic with restricted differentiation
ability. Table 1. Production of factor
dependent and independent cell lines from long-term marrow cultures
a 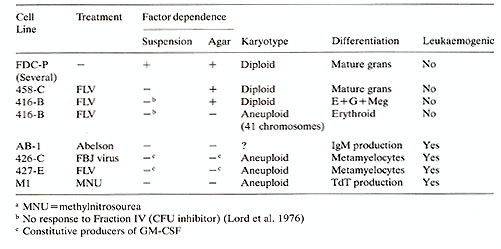
This work was supported by the Medical Research Council and the
Cancer Research Campaign. References - Dexter TM, Teich NM (1979) Modification of the proliferative and differentiation capacity of stem cells following treatment with chemical and vival leukaemogens. In: Neth R, Galls RC, Hofschneider PH Mannweiler K (eds) Modern trends in human leukaemia III. Springer, Berlin Heidelberg New York, pp 223-229 - Dexter TM, Allen TD, Lajtha LG (1977a) Conditions controlling the proliferation of hemopoietic stem cells in vitro. J Cell Physiol 91 :335-344 - Dexter TM, Scott D, Teich NM (1977b) Infection of bone marrow cell proliferation, differentiation and leukaemogenic capacity. Cell 12: 355-364 - Dexter TM, Allen TD, Lajtha LG , Krizsa F, Testa NG, Moore MAS (1978). In vitro analysis of self-renewal and committment of hemopoietic stem cells. In: Clarkson B, Marks PA, Till JE (eds) Differentiation of normal and neoplastic hemopoietic cells. Cold Spring Harbor Press, New York, pp 63-80 - Dexter TM, Allen TD, Scott D, Teich NM ( 1979) Isolation and characterisation of a bipotential hemopoietic cell line. Nature 277:471-474 - Dexter TM, Spooncer E, Toksoz D, La.jtha LG (to be published a) The role of cells and their products in the regulation of in vitro stem cell proliferation and granulocyte development. J Supramol Struct -Dexter TM, Allen TD, Teich NM (to be published b) Production of 'normal' stem cell lines following treatment of long-term marrow cultures with Friend murine leukaemia viruses. - Dexter TM, Garland J, Scott D, Scolnick E, Metcalf D (1980c) Growth of factor dependent hemopoietic precursor cell lines. J Exp Med 152: 1036-1047 - Fibach E, Landau T, Sachs L (1972) Normal differentiation of myeloid leukaemic cells induced by a differentiation inducing protein. Nature 237: 276-279 - Greenberger JS, Gans PJ, Davisson PB, Maloney WC ( 1979) In vitro induction of continuous acute promyelocyte leukaemia cell lines by Friend or Abelson murine leukaemia viruses. Blood 53 :987 Ichikawa Y (1969) Differentiation of a cell line of myeloid leukaemia. J Cell Physiol 74:223-234 - Lord BI, Mari KJ, Wright EG, Lajtha LG (1976) An inhibitor of stem cell proliferation in normal bone marrow. Br J Haematol, 34:441-445 - Lord BI, Mori KJ, Wright EG, Lajtha LG (1977) A stimulator of stem cell proliferation in regenerating bone marrow. Biamed Express (Paris) 27:223-226 - Metcalf D (1977) Hemaopoietic colonies. Springer, Berlin Heidelberg New York, p 277 - Metcalf D, Moore MAS, Warner NL (1969) Colony formation in vitro by myelomonocytic leukaemic cells. J Natl Cancer Inst 43:983-1001 - Teich NM, Dexter TM (1978) Effects af murine leukaemia virus infection on differentiation of hemapoietic cells in vitro. In : Clarkson B, Marks PA, Till JE (eds) Differentiation of normal and neoplastic hemapoietic ce1ls. Cold Spring Harbor Press, New York, pp 657-670 -Teich NM, Dexter TM (1979) Interaction between murine leukaemia viruses and differentiating hemopoietic cells. In: Oncogenic viruses and host cell genes. Academic Press, New York, pp 263-276 - Teich N, Boss M, Dexter TM ( 1979) Infection of mouse bone marrow cells with Abelson murine leukaemia virus and establishment of producer cell lines. In: Neth R, Calla RC, Hofschneider PH, Mannweiler K (eds) Modern trends in human leukaemia III. Springer, Berlin Heidelberg New York, pp 487-490 - Testa NG, Dexter TM, Scott D, Teich NM (1980) Malignant myelomonocytic cells after in vitro infection of marrow cells with Friend leukaemia virus. Br J Cancer 41:33-39 |