|
1 Department of Experimental
Haematology, Paterson Laboratories, Christie Hospital & Holt Radium
Institute, Withington, Manchester M20 9BX, England A. Introduction Haemopoietic stem cells are derived early in embryogenesis, are
relatively few in number, but persist throughout adult life by virtue
of their ability to undergo self-renewal. This ability to undergo
self-renewal is a characteristic and essential property of stem
cells: in the absence of self-renewal the haemopoietic system would
rapidly decline, while excessive and persistent self-renewal (in
the absence of death or differentiation) would lead to a dramatic
reduction in the production ofmature cells and an increase in primitive
cells, i.e. a leukaemia [24, 35]. It follows, then, that an investigation
of self-renewal and differentiation is an over-riding problem in
the understanding of growth control in normal tissues as well as
the lack of growth control which occurs during tumourigenesis. In
this context, the role of tissue and celllineage-restricted growth
factors and of oncogenes (and their products) is assuming more and
more importance [ 16, 48]. In this communication, the role of one
such growth factor (haemopoietic cell growth factor), the stromal
cell milieu, and the src oncogene are discussed in relation to self-renewal
and differentiation ofhaemopoietic cells. B. The Role of the Marrow Stroma in Haemopoiesis Haemopoietic cell deveJopment in the adult occurs mainly in the bone marrow, where the developing blood cells are found in intimate association with a stromal cell network [26, 49]. Evidence indicates that the stromal cells supply the extracellular matrix and cell-cell interactions necessary for the proliferation of stem cells [40] as well as their differentiation and development into the various myeloid lineages. For example, the stromal cells present in long-term marrow cultures produce factors which specifically (and reversibly) stimulate or inhibit DNA synthesis in CFU-S [46, 53, 54]: thus, changes in the respective concentrations of these opposing activities act to modulate proliferative activity of the stem cells. However, within the long-term cultures (and in marrow in vivo), the population size of stem cells is strictly controlled, such that in normal steady state conditions the number of CFU-S remains fairly constant [45]. This control can operate at two levels. First, at the level of stem cell proliferation where an increased demand for CFU-S (following partial ablation with cytotoxic chemicals) is met by increased proliferative activity [ 19, 52] (associated with increased production of the CFU-S "stimulator" alluded to earlier). Second, at the level of stem cell differentiation, a process which is an enigma at present, but which generates a series of lineage-restricted progenitor cells such as granulocyte/ macrophage colony-forming cells (GMCFC), erythroid colony forming cells (BFU-E) and so on. Most, ifnot all, of these committed progenitor cells can be recognised by their ability to undergo clonal expansion in soft gel systems to produce colonies containing mature cells of the appropriate cell lineages [ 13, 28]. The development of the cells within these soft gel sycstems requires the continuous presence of appropriate growth factors such as granulocyte/macrophage colony-stillulating factor (GM-CSF), burst-promoting activity (BPA), etc. However, the development of the progenitor cells in long-term marrow cultures occurs in association with the marrow stromal cells and does not require the presence of added stimulatory molecules [12,50]. Indeed, it is difficult to detect the presence of molecules such as GM-CSF or BPA in medium conditioned by the growth of marrow stromal cells in vitro, and for this reason we believe that such molecules are normally produced by the stromal cells and stimulate progenitor cells locally, where an effective concentration of the growth factor can be maintained [14, 18,37,51]. In other words, the haemopoietic system may be best viewed as consisting of a series of stromal cell niches facilitating stem cell proliferation/ differentiation or promoting progenitor cell development. This view is supported by our observation that specific stromal cell-haellopoietic cell interactions occur during erythroid cell development and that similar but different stromal cell-haellopoietic cell interactions occur during granulocyte development [1,2,11]. The net result of these complex interactions is the maintenance of homeostasis such that for everyone CFU-S there are about ten progenitor cells and for everyone GMCFC, there are between 500 and 1000 mature cells being produced. These ratios are highly consistent and are found both in marrow in vivo and in long-term cultures in vitro. Obviously, an increase or a decrease in the self-renewal probability of the stem cells may have a profound effect upon the production of mature cells -leading either to an aplasia or a hyperplasia.
Several haemopoietic growth factors have now been purified to homogeneity, partial amino acid sequence determined and, in at least two cases, molecularly cloned [8, 10, 20, 29, 33, 42, 55] and A. W. Burgess, personal communication). One of these growth factors is GM-CSF, purified from mouse lung cell-conditioned medium. This factor stimulates the development of GM-CFC to produce colonies in vitro containing neutrophils and macrophages [8]. In this case, however, little or no self-renewal occurs: after several days' growth, the GM-CFC give rise to colonies containing only mature cells [4, 7, 30]. Because of this, it was once thought that GM-CSF was a lineage-restricted regulatory molecule. Recently, however, it has been shown that multipotential stem cells can also respond to this molecule, which acts as a proliferative signal as well as facilitating differentiation of the multi potential cells to lineage-restricted progenitor cells [31]. Thus, GM-CSF may have a wider range of "target" cells than initially thought. Despite this, there is no evidence that GM-CSF is acting as a selfrenewal stimulus for multipotential cells or GM-CFC; rather, it appears to be involved in facilitating development of the stem and progenitor cells in the absence of self-renewal. A similar role can also be ascribed to the molecule which selectively stimulates granulocyte development from the GMCFC [29, 33] (so-called granulocyte colonystimulating factor, or G-CSF) and to the molecule which preferentially still ulates macrophage development from the GMCFC [42] (macrophage colony- stimulating factor, M-CSF or CSF-l ). Similarly, the hormone erythropoietin (which acts on the terminal stages of erythropoiesis) has so far not been shown to act as a self-renewal-inducing molecule (E. Spooncer, unpublished work). Consequently, the role oflineage-restricted regulatory molecules seems to be one of facilitating development rather than self- renewal. In contrast to this is the action of the molecule which we have recently purified to homogeneity from WEHI-3 cell-conditioned medium. This molecule, which we term haemopoietic cell growth factor [ 4] (or HCGF) is a glycoprotein of 25 kilo daltons with a polypeptide core of 15 kilodaltons, and is almost certainly the same molecule as that known as IL-3, BPA, PSF, mast cell growth factor or multi-CSF [9, 22, 23, 32, 36, 56]. The widespread interest in this molecule arises from the observation that it can "immortalise" granulocyte precursor cells and mast cells, i.e. it will allow such cells to selfrenew continuously in vitro [15, 21, 44]. It will also promote proliferation and self-renewal of CFU-S in short-term liquid culture [27, 47] and L. Dorssers, personal communication), as well as proliferation and development of multipotential stem cells and the various types of myeloid committed progenitor cells [4]. In other words, HCGF appears to be a "master control" molecule influencing the earliest stages of haemopoiesis. This does not mean that the lineage-restricted molecules (such as CSF-l and erythropoietin) do not have a role to play in haemopoietic cell development. Rather, they may best be seen as the "fine tuners" in the system, modulating cell production to meet the demand existing at any one time. Of major interest is that, unlike the molecules such as GM-CSF, M-CSF, G-CSF and erythropoietin, HCGF cannot be detected in the serum of mice, although it is readily produced in vitro from lectin-stimulated T cells [9]. Our proposition is that the in vivo production of this molecule is probably very stringently controlled, being normally produced only in areas where self-renewal is required. Obviously, excessive or systemic production of HCGF may be expected to have a dramatic effect on haemopoiesis. Indeed, it could be that the WEHI3 leukaemic cells (a potent source of HCGF) are leukaemic by virtue of their ability to produce HCGF and are thus "autostiillulated" for self-renewal.
From the previous discussion, it is clear that the maintenance of homeostasis requires a delicate balance between cell-cell interactions and diffusible regulatory molecules (growth factors). The products of certain oncogenes, i.e. the oncogenes associated with leukaemic transformation such as abl, myc, myb, erb, presumably alter this balance in such a way that increased self-renewal occurs at the expense of differentiation [6]. How they do this is unclear, although the evolutionary conservation of cellular oncogenes suggests an important role for their products in normal cell growth and differentiation [ 17,38,43]. Recently, we have been investigating the effects of the src oncogene (which codes for a product called pp60src), an oncogene which is associated with the production of sarcomas in chickens [5]. Using molecular recombination techniques, the avian src has been placed under the influence of an amphotropic murine virus promoter sequence [3], and this virus, src (MoMuL V), has been used to infect murine long-term marrow cultures [7,41]. The effect was dramatic.
Little change was seen in the gross morphology or organisation of the stromal elements for several weeks. However, the cultures were infected with the src (MoMuLV) between 5 and 8 weeks after initiation of the cultures -a time at which little proliferative activity is occurring in the stromal cells, hence to initial lack of effect is not unexpected. However, within 8-10 weeks after infection there occurred a progressive overgrowth of "transformed" fibroblasts and macrophages which, at later times in culture, were released from the adherent layer and were found admixed with haemopoietic cells in the growth medium [7].
Following infection with src (MoMuLV) there was a progressive
increase in the concentration and in the absolute number of stem
cells (CFU-S) and committed progenitor cells (GM-CFC) released into
the growth medium. The increases in the con centration of CFU-S
and GM-CFC (which are 50-100 times the level seen in the control)
reflected an absolute increase of 10-20 times the number seen in
the corresponding control cultures. At the same time, however, the
number of mature haemopoietic cells produced fell dramatically.
In control cultures, between 400 and 1000 mature cells were present
per GM-CFC. In infected cultures, on the other hand, the level of
mature cells progressively declined to reach a level ofabout 5-10
mature cells per GMCFC [7]. In other words, src (MoMuL V) infection
of long-term marrow cultures led to an inversion of the developmental
hierarchies seen during normal haemopoiesis. The effects are schematically
described in Fig. I. 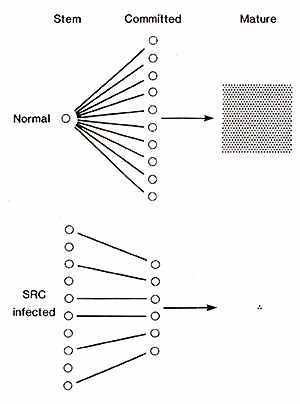
Notwithstanding the effects seen on developmen tal transi tions descri bed earlier , we repeatedly found that (irrespective of the time since infection) injection of the cells into immunocompromised or potentially lethally irradiated recipients did not lead to the development of a leukaemia [7, 41]. In fact, the spleen colonies produced from the CFU-S present in src (MoMuL V)infected cultures were normal in all aspects examined, the cells would protect mice from the effects of a lethal dose of radiation, and the reconstituted animals lived a normal life span without the emergence of leukaemic disease [41]. In these reconstituted animals a slight anaemia was occasionally observed, but no evidence of a gross block in differentiation was seen -as happened in the original src (MoMuL V)-infected long-term cultures. We conclude, therefore, that the haemopoietic cells produced in the src-infected cultures are essentially normal (at least in terms of differentiation ability) and the developmental block seen in the infected long-term culture is due primarily to an effect of src on the supportive stromal cell elements.
While the CFU-S in the src-infected cultures are apparently normal
in their ability to produce spleen colonies and to reconstitute
irradiated mice, we did detect a profound change in their self-renewal
ability. For example, when normal CFU-S are serially transferred
in vivo in irradiated recipients, they rapidly lose their ability
to produce more CFU-S or to reconstitute haemopoiesis in such mice,
after the second or third transfer [25, 34, 39]. Similarly, if normal
CFU-S are serially passaged in vitro on the irradiated stromal celllayer
of a long-term marrow culture, they likewise rapidly lose their
ability to self-renew and establish haemopoiesis [34]. CFU-S from
the src (MoMuL V)-infected cultures, however, can be repeatedly
passaged in vivo or in vitro without any apparent decline in their
ability to produce CFU-S or to establish apparently normal haemopoiesis.
Thus, these data argue that while the cells are not leukaemic, they
have 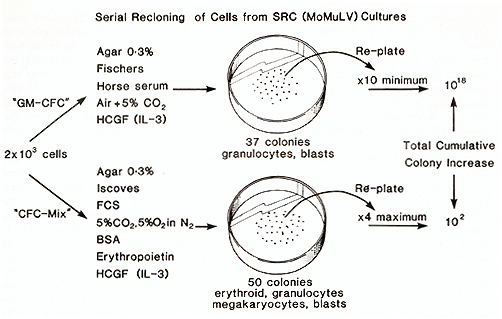
The colonies developing in the agar culture system described previously have been analysed for their developmental potential. To do this, colony cells at various transfer numbers, i.e. between one and ten serial in vitro reclonings, were plated in growth medium facilitating the development of multipotential cells [41]. We consistently found that in these conditions, between 30% and 50% of the colonies produced contained cells of more than one myeloid lineage, including erythroid cells. It seems, then, that the serial recloning ability of the cells in vitro (in a GM-CFC assay system) is associated with self-renewal of multipotential stem cells (CFC-rnix). However, when these mixed myeloid colonies were tested for their ability to undergo further serial recloning in vitro, the results were uniformly poor (Fig.2). From these data, we conclude that the self-renewal observed is in part a reflection of the differentiation pressure imposed by the different culture conditions. The results are schematically described in Fig. 2 [41].
Individual colonies developing in the GMCFC assay system (Fig. 2) were picked out and the cells resuspended in Fischer's medium supplemented with 20% horse serum and WEHI-3 cell-conditioned medium (10% vol/vol) as a source of HCGF. Of 75 individual colonies isolated in this way, all grew and formed continuously growing cell lines in liquid culture. The growth of these cells was absolutely dependent upon HCGF; in its absence, the cells died. The cell lines produced have a primitive morphology and resemble other growth factordependent cell lines (FDC-P). However, all the clones so far examined possess an ability to produce mixed myeloid colonies ( erythroid cells plus one other lineage ) when plated in soft agar. A more detailed examination of one clone FDC/Mxl has demonstrated that the cells will grow in the absence of HCGF provided they are cultured on a marrow stromal cell layer (E. Spooncer and T. M. Dexter, in preparation). In this respect, the cells are unlike other FDC-P cells, which do not retain this ability. Furthermore, the FDC/ Mx 1 cells "infiltrate" the marrow stromal cells and form foci of haemopoiesis. As these foci grow and develop, mature granulocytes and (in appropriate conditions) mature red cells are produced for many months. In this respect, therefore, the FDC/ Mx 1 cells are like normal stem cells which also have the ability to infiltrate marrow stromal cell layers and establish haemopoiesis (E. Spooncer and T. M. Dexter, to be published). Thus far, these detailed studies have only been performed with one clone of cells, FDC/Mxl. However, preliminary data suggests that this retention of stem cell character applies to all the clones isolated. Furthermore, injection of the cells in vivo has thus far not given rise to any leukaemias. Thus, the original infection of the long-term cultures with src (MoMuL V) has led to the eventual development of continuously growing, nonleukaemic, multipotential stem cell lines which require the presence of HCGF for their growth in liquid cultures, but where the HCGF can be replaced by marrow stromal cells. These cells should provide a powerful system for investigating the processes of self-renewal and differentia tion in normal haemopoiesis and leukaemogenesis.
Infection of long-term marrow cultures with other "helper" or rapidly transforming RNA tumour viruses has not led to the effects observed after infection with the src (MoMuLV) virus [6]. Therefore, it seems reasonable to conclude that the haemopoietic changes and generation of HCGFdependent multipotential stem cell lines is directly or indirectly associated with src gene expression. Indeed, previous results have clearly demonstrated high levels of pp60src, measured by its kinase activity, in both the adherent (mainly stromal) and in the nonadherent (mainly haemopoietic) cell populations of infected long-term cultures [7]. However, the continuously growing cell lines do not produce infectious src (MoMuL V) nor do they express high levels of src kinase activity (J.Wyke and A. Stoker, personal communication), although such cells clearly have an extended capacity for self-renewal compared with their normal counterparts. Thus, the role of pp60src remains an enigma. Obviously several possibilities are raised by this finding. First, that src (MoMuL V) infection of the long-term cultures "selected" for cells with a greater self-renewal ability. If this is the case, however, it is difficult to understand how the phenotype is maintained after prolonged growth in vitro in the absence of stroma. Other possible explanations include "hit and run" events, initial expression and subsequent repression of pp60src, or the integration of provirus adjacent to some important regulatory gene. However, these must be highly frequent events to account for the ease with which src (MoMuL V)-infected cultures undergo transformation and the ease with which continuously growing multipotential stem cell lines can be established from such cultures. These possibilities are being explored at present. Nonetheless, the effects observed indicate the complex events which determine self- renewal of the stem cells, the importance of the cellular environment, of the different growth factors and the effects that oncogene expression can have on these processes.
This work was supported by the Cancer Research Campaign (UK)
1. Allen TO, Dexter TM (1982) Ultrastructural aspects of erythropoietic differentiation in long-term bone marrow culture. Differentiation 21: 86-94 2. Allen TO, Dexter TM (1983) Long term bone marrow cultures: an ultrastructural review. Scan Electron Microsc 4: 1851-1866 3. Anderson SM, Scolnick EM (1983) Construction and isolation of a transforming murine retrovirus containing the src gene of Rous sarcoma virus. J Virol 46: 594-605 4. Bazill GW, Haynes M, Garland J, Dexter TM (1983) Characterization and partial purification of a haemopoietic cell growth factor in Wehi-3 cell conditioned medium. Biochem J 210:747- 759 5. Beard JW (1963) Avian virus growths and their etiologic agents. Adv Cancer Res 7: 1 -127 6. Boettiger DB, Dexter TM (1984) Long-term marrow cultures in the analysis of viral leukemogenesis. Clin Haematol13:349-370 7. Boettiger DB, Anderson S, Dexter TM (1984) Effect of src infection on long-term marrow cultures: Increased self-renewal of hemopoietic progenitor cells without leukemia. Cel136:763-773 8. Burgess AW, Camakaris J, Metcalf D (1977) Purification and properties of colony-stimulating factor from mouse lung conditioned medium. J Biol Chem 252: 1998-2003 9. Burgess AW, MetcalfD, Russel SHM, Nicola NA (1980) Granulocyte/macrophage-megakaryocyte-, eosinophil-, and erythroidcolony-stimulating factors produced by mouse spleen cells. Biochem J 185:301-314 10. Das SK, Stanley ER (1982) Structure-function studies of a colony stim ulating factor (CSF-l). J Biol Chem 257: 13579 -13684 11. Dexter TM ( 1982) Stromal cell associa ted haemopoiesis. J Cell Physiol [Suppl] 1: 87-94 12. Dexter TM, Shadduck RK (1980) The regulation of haemopoiesis in long-term bone marrow cultures. I. Role of L-cell CSF. J Cell Physiol102:279-286 13. Dexter TM, Testa NG (1980) In vitro methods in haemopoiesis and lymphopoiesis. J Immunol Methods 38: 177-190 14. Dexter TM, Allen TO, Lajtha LG (1977) Conditions controlling the proliferation of haemopoietic stem cells in vitro. J Cell Physio191 : 335-344 15. Dexter TM, Garland J, Scott D, Scolnick E, Metcalf D (1980) Growth of factor dependent haemopoietic precursor cell lines. J Exp Med 152: 1036-1047 16. Downward J, Yarden Y, Mayes F, Scrau G, Totty N, Stockwell P, Ullrich A, Sclessinger J, Waterfield MD (1984) Close similarity of epidermal growth factor receptor and V -erbB oncogene protein sequences. Nature 307: 521-527 17. Duesberg PH (1983) Retroviral transforming genes in normal cells. Nature 304:219-226 18. Eliason JF, Dexter TM, Testa N (1982) The regulation of hemopoiesis in 1ong-term bone marrow cultures III. The role of burstpromoting activity. Exp Hemat 10:444-450 19. Frindel E, Croizat H, Vassart F (1976) Stim ulating factors liberated by treated bone marrow: I n vitro effect on CFU kinetics. Exp Hemat 4: 56-61 20. Fung MC, Hapel AJ, Ymer S, Cohen DR, Johnson RM, Campbell HD, Young IG ( 1984) Molecular cloning of cDNA for murine interleukin-3. Nature 307: 233-237 21. Greenberger JS, Eckner RJ, Sakakeeny M, Marks P, Reid D, Nabel G, Hapel A, IhIe JN, Humphries KC (1983) Interleukin 3-dependent hematopoietic progenitor cell lines. Fed Proc 42:2762-2771 22. I hIe JN, Keller J, Oroszlan S, Henderson LE, Copeland TD, Fitch F, Prystowsky MB, Goldwasser E, Schrader JW, Palaszynski E, Dy M, Lebel B (1983) Biologic properties of homogeneous interleukin 3. J Immunol 131 : 282-287 23. Iscove NN, Roitch CA, Williams N, Guilbert LJ (1982) Molecules stimulating early red cell, granulocyte/macrophage, and megakaryocyte precursors in culture: similarity in size, hydrophobicity and charge. J Cell Physiol [Suppl] I: 23-30 24. Lajtha LG ( 1979) Stem cell concepts. Differ entiation 4: 23-34 25. Lajtha LG, Schofield R (1971) Regulation of stem cell renewal and differentiation: possible significance in ageing. In: Strehler BL (ed) Advances in gerontological research. Academic, New York, pp 131146 26. Lictman MA ( 1981) The ultrastructure of the hemopoietic environment of the marrow: A review. Exp Hematol 9: 391-410 27. Lowenberg B, Dicke KA (1977) Induction of proliferation of haemopoietic stem cells in culture. Exp HematoI5:314-331 28. Metcalf D (1977) Hemopoietic colonies. Springer, Berlin Heidelberg New York, p 227 29. MetcalfD, Nicola NA (1983) Proliferative effects of purified granulocyte colony-stimulating factor (G-CSF) on normal mouse haemopoietic cell. J Cell Physiol 116: 198- 206 30. Metcalf D, Johnson GR, Mandel TE (1979) Colony formation in agar by multipotential hemopoietic cells. J Cell Physiol 98: 40 1-424 31. MetcalfD, Johnson GR, Burgess AW (1980) Direct stimulation by purified GM-CSF of the proliferation ofmultipotential and erythroid progenitor cells. Blood 55: 138-147 32. Nabel G, Galli SJ, Dvorak AM, Dvorak HF, Cantor H (1981) Inducer T-Iymphocytes stimulate a factor that stimulates proliferation of cloned mast cells. Nature 291 :332 -334 33. Nicola NA, Metcalf D, Matsumoto M, Johnson GR (1983) Purification of a factor inducing differentiation in murine myelomonocyte leukemia cells. J BioI Chem 258:9017-9023 34. Schofield R, Dexter TM (1984) Studies on the self-renewal ability of CFU-S which have been serially transferred in long-term culture or in viva. Leuk Res (in press) 35. Schofield R, Lajtha LG (1983) Determi nation of the probability of self-renewal in haemopoietic stem cells: a puzzle. Blood Cells 9:467-473 (1983) 36. Schrader JW, Clark-Lewis I (1982) AT -cell derived factor stimulating multipotential hemopoietic stem cells: molecular weight and distinction from T -cell growth factor and T-cell derived granulocyte macrophage colony-stimulating factor. J Immunol 129: 30-35 37. Shadduck RK, Waheed A, Greenberger JS, Dexter TM (1983) Production of colonystimulating factor in long-term bone-marrow cultures. J Cell Physioll14: 88-92 38. Shilo B, Weinberg RA (1981) DNA sequences homologous to vertebrate oncogenes are conserved in Drosophila melanogaster. Proc Natl Acad Sci USA 78:6789-6792 39. Siminovitch L, Till JE, McCulloch EA (1964) Decline in colony-forming ability of marrow cells subjected to serial transplantation into irradiated mice. J Cell Comp Physiol 64: 23 -31 40. Spooncer E, Gallagher JT, Krizsa F, Dexter TM (1983) Regulation of haemopoiesis in long term bone marrow cultures. IV. Glycosaminoglycan synthesis and the stimulation of haemopoiesis by ß-D-xylosides. J Cell BioI 96:510-514 41. Spooncer E, Boettiger DB, Dexter TM (1984) Continuous in vitro generation of clonogenic multipotential stem cells from src infected cultures. Nature (in press) 42. Stanley ER (1979) Colony stimulating factor (CSF) radioimmunoassay: Detection of a CSF subclass stimulating macrophage production. Proc Natl Acad Sci USA 76:2969 -2973 43. Tatchell K, Chaleff DT, Defeo-Jones D, Scolnick EM (1984) Requirement of either of a pair of ras-related genes of saccharomyces cerevisiae for spore viability. Nature 309: 523-527 44. Tertian G, Yung TP, Guy-Grand D, Moore MAS ( 1981) Long term in vitro culture of murine mast cells. I. Description of a growth factor-dependent culture technique. J Immuno1127: 788- 794 45. Toksoz D (1980) Ph.D. Thesis, University of Manchester 46. Toksoz D, Dexter TM, Lord BI, Wright EG, Lajtha LG (1980) The regulation of hemo poiesis in long-term bone marrow cultures. II. Stimulation and inhibition of stem cell proliferation. Blood 55:931-936 47. Wagemaker G, Peters MF (1978) Effects of human leukocyte conditioned medium on mouse haemopoietic progenitor cells. Cell Tissue Kinet II :45-56 48. Waterfield MD, Scrace GT, Whittle N, Stroobart D, Johnsson A, Wasterson A, Westermark B, Heldin CM, Huang JS, Duel TF ( 1983) Platelet derived growth factor is structurally related to the putative transforming protein of p28 sis of Simian sarcoma virus. Nature 304: 35-39 49. Weiss l (1981) Haematopoiesis in mammalian bone marrow. In: Porter R, Whelan J ( eds ) Cell in teractions in haellopoietic dif ferentiation. Ciba Foundation Symposium 84. Pitman Medical, Tunbridge Wells, pp I -4 50. Williams N, Burgess A W (1980) The effect of mouse lung granulocyte macrophage colony stimulating factors and other colony stimulating activities on the proliferation and differentiation of murine bone marrow cells in long-term cultures. J Cell Physiol 102 : 287 -295 51. Williams N, Jackson H, Rabellino E (1977) Proliferation and differentiation of normal granulopoietic cells in continuous bone marrow cultures. J Cell Physiol 93: 435-440 52. Wright EG, Lord BI ( 1977) Regulation of CFU-S proliferation by locally produced endogenous factors. Biomed Express 27: 215 -218 53. Wright EG, Garland JM, Lord BI (1980) Specific inhibition of haemopoietic stem cell proliferation: characterization of inhibitor producing cells. leuk Res 4:537-545 54. Wright EG, Ali AM, Riches AC, Lord BI (1982) Stimulation of haemopoietic stem cell proliferation: characteristics of still ulator producing cells. leuk Res 6: 531-539 55. Yokota T, lee F, Rennick D, Hall C, Arai N, Mosmann T, Nabel G, Cantor H, Arai K (1984) Isolation and characterization of a full-length cDNA for mast cell growth factor from a mouse T -cell clone: expression in monkey cells. Proc Nat Acad Si USA 81: 1070-1074 56. Yung YP, Eger R, Tertian G, Moore MAS (1981) long-term in vitro culture of murine mast cells. II. Purification of a mast cell growth factor and its dissociation from TCGF. J I rnrnunol127:794-799 |