|
Department of Microbiology. University of Louvain. UCL 30.58. B 1200 Brussels. Belgium
Bacterial plasmids have been known for more than twenty years and, during this period. one witnessed an extraordinary spread of those extra-chromosomal DNA elements. Resistance plasm ids were first identified in Enterobacteriaceae but, later on. have been found in almost every pathogenic bacterial species, including more recently Hemophilus influenzae and Neisseria gonorrheae. Plasmids, however. do not only contribute to antibiotic resistance. They can code for many other properties, including heavy metal resistance, production ofa cholera-like enterotoxin, tumorigenicity in plants and the capacity to catabolize various substrates. Some plasm ids have been found to code for lactose fermentation and this type of plasmid also happens to be of medical concern. Their presence can indeed obscure the detection of pathogens such as Salmonella and Yersinia since the capacity to ferment lactose is the major key in the sorting of the Enterobacteriaceae. Plasmids consist of covalently closed circular DNA and their essential feature is a replication region that ensures the propagation of the entire structure. Some plasmids also contain an operon promoting the transfer of the plasmid itself from one bacteria to another one.
In 1974. Hedges and Jacob observed that the ampicillin resistance
determinant of a plasmid called RP4 could be translocated to the
bacterial chromosome and various other plasmids genetically unrelated.
From one of these derivative plasmids, they could subsequently translocate
the ß-lactamase gene to a third plasmid. After transposition, the
increase in molecular weight was similar in every instance. These
authors concluded that a DNA sequence, including the ß-lactamase
gene, had a specific transposition mechanism and they called it
a transposon (now Tn I). Later on, Berg et al. (1975) observed the
transposition of the kanamycin resistance genes from two different
resistance plasm ids to bacteriophage gamma (Tn5 and Tn6). Gottesman
and Rosner (1975) made similar observations with a determinant for
chloramphenicol resistance: initially detected on a resistance plasmid
and. later. transferred to phage Pl, it could subsequently be translocated
from Pl to bacteriophage gamma (Tn9). Still in the same year, Kleckner
et al. (1975) reported that agenetic elemen t. 8,3 Kb long, carrying
the tetracycline resistance gene was also capable of transloca tion
(Tn 10). Recently. we showed that genes coding for lactose fermentation
can jump from pGC 1. a plasmid that originated in Yersinia enterocolitica,
to RPI, a resistance plasmid that originated in Pseudomonas aeruginosa
(Cornelis et al., 1978) (Fig. 1 ). This lactose transposon that
alters the biochemical phenotype of its host is 16,6 Kb long and
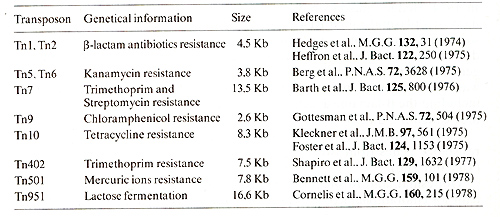
has been given the number 951. Some transposons are listed in Table
1. 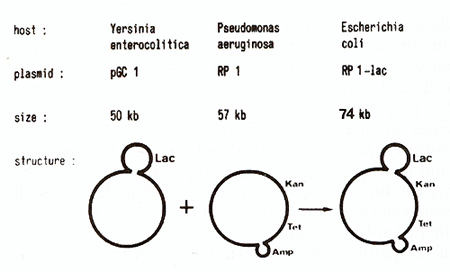 Fig. I. Schematic representation of the transposition ofTn951. carrying a lactosc operon. from pGCI to RPl. RPl confers resistance to kanamycine (Km). tetracycline (Tc) and ampicillin (Amp). The latter gene is part of another transposon (Tn I)
As suggested by Hedges and Jacob ( 1974 ). transposons are specific DNA sequences of a precise length. The size of any replicon suffering transposition does increase and the increment is constant for a given transposon. Transposition does not require extended homology between the two replicons exchanging the transposon. This is shown in Table 2 for pGCl (lac+, tra ts, inc unknown) and RPI (Amp, kan, tet, tra, inc P), the two plasmids exchanging Tn95l. Thus, transposition cannot involve ordinary recombination occuring after physical breaking and reciprocal exchange of DNA. Accordingly. in some instances at least, transposition turns out to be independant of the rec A gene product, the bacterial function involved in such recombination process (Rubens et al., 1976). Table 2. DNA-DNA hybridization between
pGCI and RPI  Transposition is therefore considered as some kind of "illegitimate" recombination involving the termini. The known transposons can jump on a great number of targets such as different plasmids, bacteriophages and bacterial chromosomes. There seems however to be some kind of specificity: Tnl for instance was found by Hedges and Jacob ( 1974) to transpose from RP4 to three different plasmids but not on a fourth one. Bennett and Richmond (1976) also observed that the transposition frequencies of Tn 1 could vary 10000 times according to the chosen target. Kretschmer et al. (1977) observed the same phenomenon with Tn3. Inserted transposons can readily be detected on small replicons such as plasmids and bacteriophages by electron microscopy. When a mixture of two different DNAs sharing homologous regions is denatured and allowed to reanneal slowly. a number of hybrid molecules are generated. If such "heteroduplex" molecules are constructed between a given plasmid and its own derivative carrying a transposon, the structure observed in electron microscopy will be a circular double stranded plasmid carrying a single stranded loop corresponding to the transposon. This kind of analysis clearly demonstrates that transposons are inserted without any loss of DNA from the recipient plasmid. Moreover. heteroduplex analysis will indicate whether the insertions on two independantly isolated molecules occur at the same site or at two different sites. If the transposon maps at two different loci on the target plasmid, 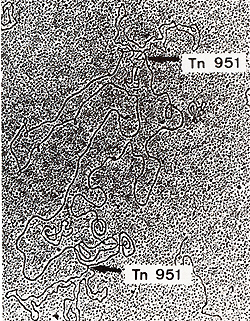
heteroduplex molecules will consist of a double stranded circular
molecule having the length of the parent plasmid and two single
stranded loops corresponding to the transposon (Fig. 2). The distance
between the two insertion sites can be measured, but this type of
analysis however doesn't allow to localize the insertion sites on
the plasmid map, at least without any refinements. This kind of
information can be inferred from experiments using restriction endonucleases.
These nucleases cleave DNA at very specific sites and thus generate
defined fragments from a given replicon. These fragments are subsequently
resolved by agarose gel electrophoresis. Partial digestions and
double digestions using simultaneously two different nucleases allow
to align these fragments and to draw a map, referred to as the physical
map. Insertion of a transposon into a plasmid will either introduce
new cleavage sites, generating new bands. or specifically increase
the length of a given fragment. If one knows the physical map of
the target plasmid, it is possible to map the transposition sites
and even to determine the orientation of the transposon, making
use of the restriction nucleases. Electron microscopy and restriction
nucleases have been used to map the insertion sites of various transposons
on a number of targets. Clearly, the number of insertion sites is
usually very high: we mapped eight insertions of Tn951 on RPl and
found eight different loci (Fig. 3). Heffron and co-workers ( 1975
a) could detect at least 19 sites for Tn2 on an 8 Kb long target
plasmid! Considering that transposition consists of the insertion
of DNA sequences of a certain length into a genome, it is not surprising
that it sometimes leads to the inactivation of genes. Plasmid pGC530
is an RPl : Tn951 derivative where the insertion was found to map
at coordinate 13 Kb, i.e. within the 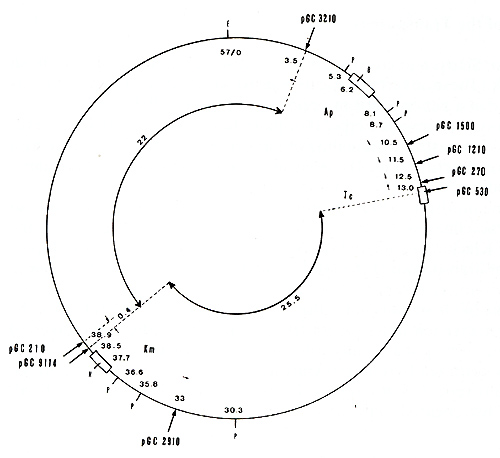 Fig.3. Physical map of RPl showing the 8 insertion sites ofTn9Sl. pGC32l0. pGClS00. pGC1210. pGC270. pGCS30, pGC2910. pGC9114 and pGC2l0 are the 8 RPl : :Tn9S1 derivatives. E = Eco Rl endonuclease cleavage site: B = Barn HI endonuclease cleavage site: P = Pst 1 endonuclease cleavage site: H = Hind III endonuclease cleavage site: Ap = ampicillin resistance gene: Tc = tetracycline resistance gene: Km = kanamycin resistance gene. Coordinates are in kilohases. The inner lines refer to distances measured in heteroduplex analysis tetracycline resistance gene (see Fig. 3). In accordance with
our physical mapping, pGC530 does no more confer resistance to tetracycline.
Insertion of a transposon not only abolishes the function of the
gene into which the element lands but it can also be polar, affecting
the expression of the genes located downstream on the transcriptional
wave (Kleckner et al., 1975). For instance. Tn2 can land in the
sulfonamide resistance gene of a plasmid and this inhibits the expression
of the neighbouring streptomycin resistance gene. Since transposons
behave like mutagens and since their insertion can readily be physically
mapped, they appear as valuable tools for the genetical analysis
ofplasmids. Tn7 has recently been used to analyse the transfer genes
of the plasmid RP4 (Earth et al., 1978). Transposons can sometimes
be excised from a replicon where they have been inserted but the
excision frequency is usually lower than the insertion frequency.
Moreover, the process is often not precise. In our hands, a strain
carrying pGC530 does not revert to tetracycline resistance, indicating
that Tn951 does not excise accurately at a detectable frequency.
D. Structure of the Transposons Five years ago, Sharp and co-workers
( 1973) discovered inverted repeats on plasmid DNA. These inverted
repeats consist of identical DNA segments occuring twice in a genome
but in opposite orientation. When a DNA fragment containing such
inverted repeats or palindromes is submitted to heteroduplex analysis.
intrastrand annealing of these fragments takes place giving a mushroom
like structure (Fig. 4), consisting ofa double stranded stem and
a single stranded loop. Later on, the mushroom like structure observed
by Sharp et al. turned out to contain tetracycline resistance genes,
in the loop. Moreover, the tetracycline transposon ofKleckner et
al. (1975) gave the same structure in heteroduplex analysis. Other
transposons were also found to consist of a DNA sequence flanked
by two inverted repeats. For instance, Tn 1 is flanked by a repeated
sequence about 140 base pairs long (Rubens et al" 1976) and Tn5
is flanked by a repeated sequence ten times longer (Berg et al.,
1975), These two inverted repeats do not correspond to any known
Insertion Sequence. Tn9, on the other hand, is bordered by the well
known sequence ISl (Starlinger and Saedler. 1976) but repeated in
tandem. Transposons are thus, often, if not always, flanked by repeated
sequences. 
Although a number ofmodels have been proposed, we have, so far, no clear understanding of the transposition mechanism. Experimental data from Heffron et al. ( 1977) suggest that a central region and the termini of Tn2 are required for transposition. The most attractive interpretation of their data is that the terminal sequence of the transposon serve a structural role while the central region encodes an enzyme required for transposition. Apart from that. it is not yet clear whether the process requires the recognition of a special sequence on the target.
The discovery of transposable elements seems to account significantly for the structural and genetic diversity of plasmids. It has been known for long indeed that there is no relation between the genes carried by a plasmid and the type ofplasmid. For instance, the TEM ß-lactamase can be coded by plasmids that belong to. at least, 14 different incompatibility groups (Matthew and Hedges. 1976). It is very tempting to believe that the spread of Tn land Tn2 accounts for this situation. In accordance with this hypothesis, Heffron et al. (1975b) showed that. indeed. a great variety of different plasm ids encoding the TEM lactamase contain a 4,5 Kb long sequence in common, while the TEM gene itself is much smaller. The evolution of plasmids would thus essentially be due to terminus-sitespecific recombination. Plasmids themselves seem to consist of two parts: one is essential for replication and transfer of the plasmid, while the other is a succession of transposons (and defective transposons?) acquired and lost during the history of the plasmid. In other words, plasm ids would be nothing else than genetic carriers whose function is to replicate and propagate transposons. So far, little is known about the origin of the transposons carried genes. For instance. the TEM ß-lactamase coded by Tnl and Tn2 appears different from all the known chromosomal ß-lactamases. For the lactose fermentation genes, however, we observed that the Tn951 coded operon is identical to the E. coli chromosomal lactose operon. In heteroduplex analysis. we detected 5,6 Kb homology and this length accounts for the genes encoding the repressor, the ß-galactosidase and the permease (Fig. 5) (Cornelis et al.. 1978). This indicates that, at least in some case, a transposon colded and a chromosomal (to date non transposable) operon may evolve from a common ancestor. 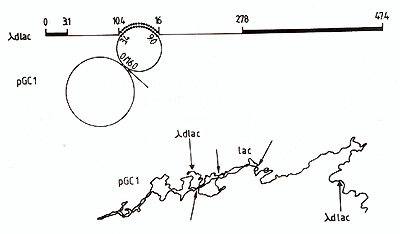 Fig.5. Diagram of an heteroduplex molecule showing the 5.6 Kb homology between the E. coli chromosomal lactose operon (here carried by gamma hdlac) and the Tn9511ac operon. here carried by pGCl
Barth. P.T.. Datta. N.. Hedges. R.W.. Grinter. N.J.: Transposition of a deoxyribonucleic acid encoding trimethoprim and streptomycin resistances from R483 to other replicons. J. Bacteriol. 125, 800-810 ( 1976) Barth. P.T.. Grinter. N. J.. Bradley. D. E.: Conjugal transfer system of plasmid RP4: Analysis by transposon 7 insertion. J. Bacteriol.133, 43-52 (1978) Bennett. P. M.. Grinsted. J.. Choi. C. L.. Richmond. M. H.: Characterization ofTn501. a transposon determining resistance to mercuric ions. Molec. Gen. Genet. 159, 101-106 ( 1978) Bennett. P. M.. Richmond. M. H.: Translocation ofa discrete piece ofdeoxyribonucleic acid carry ing an amp gene between replicons in Escherichia coli. J. Bacteriol. 126, 1-6 ( 1976 ) Berg. D.N.. Davies. J.. Allet. B.. Rochaix. J.-D.: Transposition of R factor genes to bacterio phage gamma. Proc. Natl. Acad. Sci. USA72, 3628-3632 (1975) Cornelis. G.. Ghosal. D.. Saedler. H.: Tn951 : Anew transposon carrying a lactose operon. Molec. Gen. Genet. 160,215-224 ( 1978) Foster. T.J.. Howe. T.G.B.. Richmond. K.M.V.: Translocation of the tetracycline resistance determinant from R100-1 to the E. coli KI2 chromosome. J. Bacteriol. 124, 1153-1158 ( 1975) Gottesman. M. M.. Rosner. J. L.: Acquisition of a determinant for chloramphenicol resistance by coliphage gamma. Proc. Natl. Acad. Sci. USA 72,5041-5045 ( 1975) Hedges. R.W.. Jacob. A. E.: Transposition of ampicillin resistance from RP4 to other replicons. Molec. Gen. Genet. 132, 31--40 (1974) Heffron. F.. Bedinger. P.. Champoux. J.J.. Falkow. S.: Deletions affecting the transposition ofan antibiotic resistance gene. Proc. Natl. Acad. Sci. USA74, 702-706 (1977) Heffron. F.. Rubens. C.. Falkow. S.: Translocation of a plasmid DNA sequence which mediates ampicillin resistance: Molecular nature and specificity of insertion. Proc. Natl. Acad.Sci. USA72,3623-3627(1975a) Heffron. F.. Sublett. R.. Hedges. R.W.. Jacob. A.. Falkow. S.: Origin of the TEM ß-lactamase gene found on plasmids. J. Bacteriol. 122,250-256 ( 1975 b ) Kleckner. N.. Chan. R.K.. Tye. B.-K.. Botstein. D.: Mutagenesis by insertion of a drug-resistance element carrying an inverted repetition. J. Mol. BioI. 97,561-575 ( 1975) Kretschmer. P.J.. Cohen. S.N.: Selected translocation of plasmid genes: Frequency and regional specificity of translocation of the Tn3 element. J. Bacteriol.130, 888-899 ( 1977) Matthew. M.. Hedges. R. W.: Analytical isoelectric focusing ofR factor-determined ß-Iactamases: Correlation wi th plasmid compatibility. J. Bacteriol. 125,713-718 ( 1976 ) Rubens. C.. Heffron. F.. Falkow. S.: Transposition of a plasmid deoxyribonucleic acid sequence that mediates ampicillin resistance: Independence from host rec functions and orientation of insertion. J. Bacteriol. 128,425-434 ( 1976 ) Shapiro. J. A.. Sporn. P. : Tn402: A new transposable element ldetermining trimethoprim resistance that inserts in bacteriophage gamma . J. Bacteriol. 129, 1632-1635 ( 1977) Sharp. P.A.. Cohen. S. N.. Davildson. N.: Electron microscope heteroduplex studies of sequence relations among plasmids of E. coli. II. Structure of drug resistance (R) factors and F factors. J. Mol. BioI. 75,235-255 ( 1973) Starlinger. P.. Saedler. H.: IS-elements in microorganisms. Curr.
Top. Microbiol. Immunol. 75, 111-152(1976) |