|
1 Cattedra di Oncologia, Universita di Padova,
via Gattamelata 64, 35128 Padova, Italy
2 Istituto di lstologia ed Embriologia Generale, Universita di Torino,
Corso M. D'Azeglio, 10126 Torino, Italy
3 Henry KaplanAward for the best poster Virological Session
A.Introduction
Abelson murine leukemia virus (A-MuLV) is a replication-defective
retrovirus capable of rapidly inducing leukemia in mice as well
as of transforming in vitro mouse bone marrow cells and fIbroblasts
[1, 2, 14]; the A-MuL V was derived by passage in vivo of the replication-competent
Moloney leukemia virus (M-MuLV) and its genome codes for a single
polypeptide which is a hybrid molecule containing a portion of the
parental M-MuLV genome (gag gene) and a portion of the cellular
gene termed abl [ 18], This protein, which varies in size from 160
to 90 kilodaltons, depending on the specific A-MuL V strain, is
associated mainly with the detergent-insoluble cell fraction and
possesses a tyrosine kinase activity [ 4, 15, 19], In this regard,
the A-MuLV protein resembles the pp60src encoded by the Rous sarcoma
virus (RSV) [10] and the protein kinases encoded by other retroviruses
such as the feline sarcoma virus (FeSV) [3, 17] and the Fujinami
sarcoma virus (FuSV) [9], The fact that the RSV pp60,\'rc is linked
with specialized cellular areas such as cell-cell junctions and
adhesion plaques has suggested that the alteration of these structures
is involved in the origin of the transformed phenotype [12, 13]
(Marchisio et al., Exp. Cell Res., in press). However, it has also
been demonstrated that, in the same cell type, different cellular
substrates are phosphorylated at tyrosine residues by diverse protein
kinases (Di Renzo et al., submitted). Accordingly, we found it of
interest to study the cellular substrates of the A-MuL V protein
kinase in cells having different cytoskeletal architecture and adhesion
properties such as fibroblasts and lymphocytes.
B.Materials and Methods
I. Cell Lines
TA-3 and TA-4, non-B lymphoma cell lines were established from
two independent thymic lymphomas induced by inoculating intrathymically
(i.t.) newborn BALB/c mice with the complex A-MuLV(MMuLV). MZ-5,
a pre-B lymphoma cell line, was established from a splenic lymphoma
induced by inoculating the complex A-MuLV(M-MuLV) subcutaneously
(s.c.) in newborn BALB/c micc. ABC-I, a A-MuLV-transformed line
of pre-B cells, was kindly provided by Dr. Natalie Teich []6]. ANN-]
is an A-MuLV-transformed line of fibroblasts [ 14]. As control,
a T cell lymphoma line (TB-5) induced by M-MuLV in BALB/c mice was
also included in this study. Lymphoma cell lines were cultured in
complete medium consisting or Dulbecco-MEM (Gibco Europe, Glasgow,
Scotland) supplemented with L-glutamine, HEPES, 2-mercaptoethanol,
antibiotics, and 10% heat-inactivated fetal calfserum (FCS Gibco),
ANN-l fibroblasts were cultured in Dulbecco-MEM plus 10% FCS.
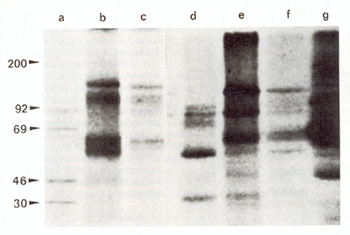
Fig. 1. Immunoprecipitation by anti-phosphotyrosine (anti-P-
Tyr) antibodies of 32p-Iabeled detergent-insoluble proteins from
A-MuLVtransformed lymphocytes and fibroblasts. Lane a molecular
weight markers; lanes b and f pre-E lymphoma cells MZ-5 and AEC-I
; lanes c and e non-E lymphoma cells TA-4 and TA-3; lane d TB-5,
M-MuLV-transformed T -cells; lane g ANN-I fibroblasts
II. Specific Antisera
Antibodies against phosphotyrosine residues (anti-P- Tyr antibodies)
were produced as previously described [6], AntiM-MuLV serum (Lot
No, 71S1161) was obtained from the Office of Program Resources and
Logistics, NCI, Bethesda, Maryland,
III. Immunoprecipitation Assay
Detergent-insoluble fractions were labeled according to Burr [5]
by incubating detergent-insoluble proteins with 32p-Iabeled y-ATP
(specific activity 5000 mCilmM, Amersham) in conditions allowing
phosphorylation catalyzed by the kinase coded by A-MuLV [4,5], After
phosphorylation, the proteins were immunoprecipitated either by
anti-P- Tyr antibodies or by anti-M-MuLV serum, as previously described
[6], After elution from protein A-Sepharose with Laemmli buffer
[II], proteins were separated by SDS-PAGE, Dried gels were exposed
to Kodak X-Omat film and processed for auforadiography.
C.Results and Discussion
The natural targets for in vivo transformation by A-MuLV are pre-B
lymphocytes [I, 2]; however, the A-MuLV, if inoculated i.t., is
also able to induce thymic lymphomas [7], Thus, in addition to ABC-l
cells we also studied some lines derived from A-MuLV induced thymic
lymphomas; these cells do not express either T cell or B cell markers,
even after stimulation with Con-A or LPS, respectively, and accordingly
were putatively defined as non-B cells, In order to investigate
whether a specific substrate for A-MuLV kinase could be detected
in different A- MuLV transformed cell types, the detergent-insoluble
cellular fraction was studied with the aid of monospecitic antibodies
directed against the phosphorylated form of protein tyrosine residues
(anti-P- Tyr antibodies), As shown in Fig, 1, two main proteins
of 70 and 120 kilodaltons were precipitated from ANN-1 fibroblasts
(Fig, 1, lane g),
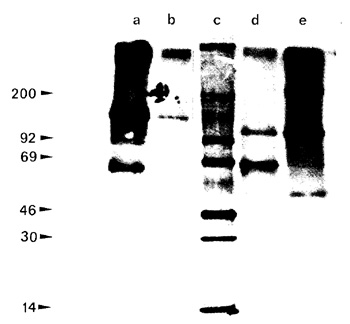
Fig.2. Immunoprecipitation of phosphotyrosine proteins
by anti M-MuLV serum and anti P-lyr antibodies in TA-3 lymphoma
cells and ANN-I fibroblasts. Lane c molecular weight markers; lanes
a and b detergcnt-insoluble traction from TA-3 lymphoma cells immunoprecipitatcd
by anti P- Tyr antibodies and anti M-MuLV serum, respectively; lanes
d and e detergent-insoluble traction from ANN-I fibroblasts immunoprecipitated
by anti-P- Tyr antibodies and anti-M-MuL V serum, respectively
whereas three different tyrosine phosphoryla ted proteins of 150,
100, and 65 kilodaltons were immunoprecipitated from pre-B and non-E
lymphoma cells (Fig. I, lanes b, c, e,f). No phosphorylated proteins
were detected in nontransformed fibroblasts and normal thymus cells
(data not shown in Fig. I). On the contrary, in the TB-5 M-MuLV
lymphoma cells, two proteins of 55 and 30 kilodaltons and an additional
one of 100 kilodaltons, comigrating with the corresponding protein
of A- MuLV lymphoma cells, were detected (Fig. I, lane d). Moreover,
in a preliminary experiment, using the anti-M-MuLV serum, which
is able to recognize the gag-coded portion of the kinase protein,
we observed only one immunoprecipitate band of 120 kilodaltons in
ANN-I fibroblasts (Fig. 2, lane e) and of 150 kilodaltons in lymphoma
cells (Fig. 2, lane b). The anti-M-MuLV serum was not able to precipitate
either the 70 kilodaltons phosphoprotein in fibroblasts (Fig. 2,
Jane d) or the 100 and 65 kilodaltons phosphoproteins in lymphoma
cells (Fig. 2. lane b), suggesting that these latter are probably
cellular substrates and not degradation products of the A-MuLV tyrosine
itself. These data seem to indicate that A-MuL V protein kinase,
under identical experimental conditions, is able to phosphorylate
at tyrosine residues different substrates in cells possessing diverse
cYtoskeletal architecture. It is known that A-MuLV-coded protein
kinase, like other transforming protein kinases, does not discriminate
for substrate phosphorylation [8, 18]; consequently, the different
pattern of phosphorylation induced by A-MuL V protein kinase in
fibroblasts and lymphocytes is probably imputable to a different
association of the tyrosine kinase with the cytoskeletal macromolecules.
Acknowledgments This work was supported by the Consiglio Nazionale
delle Ricerche, Progetto Finalizzato Oncologia, and Assoziazione
Italiana per la Ricerca suI Cancro. D.S. is a recipient of a Fondazione
Assicurazioni Generali training grant.
References
1. Abelson HT, Rabstein LS ( 1970) Lymphosarcoma. virus-induced
thymic-independent disease in mice. Cancer Res 30.2213-2222
2. Baltimore D, Rosemberg N, Wittc ON ( 1979) transformation of
immaturc lymphoid cells by Abelson murine leukcmia virus.lmmunol
Rev 48. 1-22
3. Barbacid M, Beemon K, Devarc SG ( 1980) Origin and functional
properties of the major genc product of the Snyder- Theilen strain
of feline sarcoma virus. Proc Natl Acad Sci USA77:5158-5162
4. Boss MA, Dreyfuss G, Baltimore D (1981) Localization of the Abelson
murine leukemia virus protein in a detergent-insoluble subcellular
matrix. architecture of the protein. J ViroI40:472-481
5. Burr JG, Dreyfuss G, Penman S, Buchanan JM (1981) Association
of the src gene product of Rous sarcoma virus with cytoskeletal
structures of chicken embryo fibroblasts. Proc Natl Acad Sci USA
77:3484-3488
6. Comoglio PM, DiRenzo MF, Tarone G, Giancotti FG, Naldini L, Marchisio
PC (1984) Detection of phosphotyrosine-containing proteins in the
detergent-insoluble fraction of RSV-transformed fibroblasts by azobenzene
phosphonate antibodies. EMBO J 3:483-489
7. Cook WD (1982) Rapid thymomas induced by Abelson murine leukemia
virus. Proc Natl Acad Sci USA79.2917-2921
8. Erikson RL, Collett MS, Erikson E, Purchio AF ( 1979) Evidence
that the avian sarcoma virus transforming gene product is acyclic
AMP-independent protein kinase. Proc Natl Acad Sci USA 76: 6260-6264
9. Feldman RA, Wang E, Hanafusa H (1983) Cytoplasmic localization
of the transforming protein of Fujinami sarcoma virus: saltsensitive
association with subcellular components. J Virol 45: 782- 791
10. Hunter T, Sefton BM (1980) Transforming gene product of Rous
sarcoma virus phosphorylates tyrosine. Proc Natl Acad Sci USA77.1311-1315
11. Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 277:680-685
12. Rohrschneider LR (1979) lmmunofluorescence on avian sarcoma
virus transformed cells. localization of the src gene product. Cell
16.11-14 13. Rohrschneider LR ( 1980) Adhesion plaq ues of Rous
sarcoma virus-transformed cells contain the src gene product. Proc
Natl Acad Sci USA77:3514-3518
14. Scher CD, Siegler R (1975) Direct transformation of 3T3 cells
by Abelson murine leukaemia virus. Nature 253.729- 731
15. Sefton BM, Hunter T, Raschke WC (1981) Evidence that the Abelson
virus protein functions in vivo as a protein kinase that phosphorylates
tyrosine Proc Natl Acad Sci USA78.1552-1556
16. Teich N, Boss M, Dexter TM (1979) Infection of mouse bone marrow
cells with Abelson murine leukemia virus and establishment of producer
cells. In: Neth R, Gallo RC, Hofscheider PM, Mannweiler K (eds)
Modern trends in human leukemia III. Springer, Heidelberg, pp 487-490
17. Van de Ven WJM, Reynolds FH, Stephenson JR ( 1980) The nonstructural
components of polyproteins encoded by replication-defective mammalian
transforming retroviruses are phosphorylated and have associated
protein kinase activity. Virology 101.185-197
18. Wang JYJ, Queen C, Baltimore D (1982) Expression of an Abelson
murine leukemia virus-encoded protein in Escherichia coli causes
extensive phosphorylation of tyrosine residues. J Bioi Chem 257:
13181-13 184
19. Witte ON, Rosemberg N, Paskind M, Shields A, Baltimore D ( 1978)
Identification of an Abelson murine leukemia virus-encoded protein
present in transformed fibroblasts and lymphoid cells. Proc Natl
Acad Sci USA 75.2488-2492
|